甲基化抑制劑是表觀遺傳學(xué)的載體
表觀遺傳學(xué)被定義為“不依賴于 DNA 序列的變化而是由于染色體改變所產(chǎn)生的穩(wěn)定可遺傳的表型”。DNA 甲基化,組蛋白結(jié)構(gòu)的改變以及 microRNA 對(duì)基因的調(diào)控等都是表觀遺傳信息的重要載體。其中 DNA 甲基化作為重要的表觀遺傳修飾之一,對(duì)基因組穩(wěn)定性,轉(zhuǎn)錄和發(fā)育產(chǎn)生深遠(yuǎn)影響。
甲基化到底是什么?
胞嘧啶第五碳的甲基化 (5- 甲基胞嘧啶:5mC) 是最早在真核生物中被挖掘出的甲基化類型,它也是今天我們要介紹給大家的重點(diǎn)。早在幾十年前,就有體內(nèi)外研究表明,它與轉(zhuǎn)錄抑制有關(guān)。哺乳動(dòng)物中,5mC 往往存在于 DNA 雙螺旋中的鳥(niǎo)嘌呤 (G) 的上游,即所謂的 CpG 位點(diǎn)。除了 5mC,其他的甲基化類型有:4- 甲基胞嘧啶 (4mC),5- 甲基胞嘧啶 (5mC) 和 6- 甲基腺嘌呤 (6mA)。
DNA 甲基化具有多種功能:與轉(zhuǎn)座子和基因的抑制有關(guān),但也與活躍轉(zhuǎn)錄的基因體 (gene body) 有關(guān),在某些情況下還與基因激活本身有關(guān)。
DNA 是如何出現(xiàn)表觀遺傳修飾或標(biāo)記呢?下面以哺乳動(dòng)物中 5mC 為例,揭開(kāi) DNA 甲基化修飾的神秘面紗!
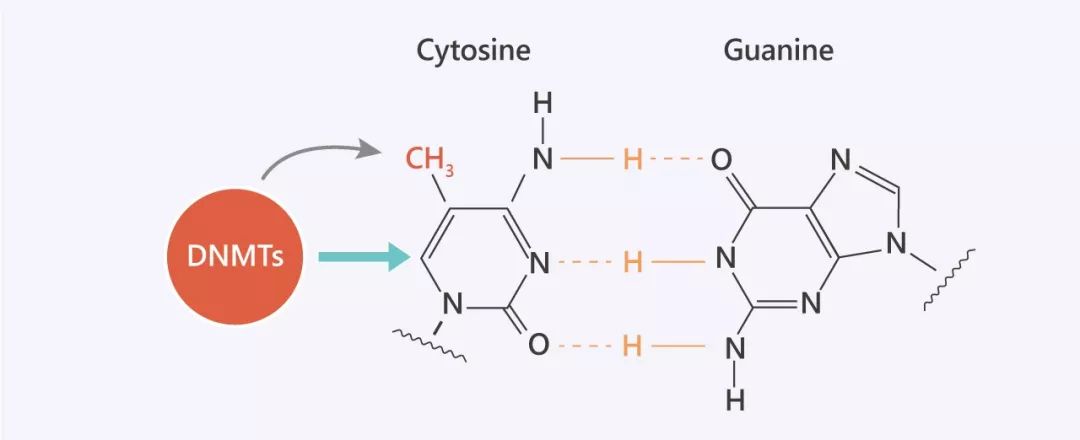
圖 1. DNA 復(fù)制過(guò)程中 CG 配對(duì)
Tip 1: CpG 是胞嘧啶 “C” 和鳥(niǎo)嘌呤 “G” 的縮寫,由磷酸酯 “p” 連接。在哺乳動(dòng)物中,胞嘧啶甲基化主要發(fā)生在 CpG 二核苷酸中。可通過(guò)對(duì)二核苷酸序列 5'CpG3' 中胞嘧啶堿基的甲基化共價(jià)修飾脊椎動(dòng)物的 DNA。
DNA 甲基化分為三個(gè)階段:甲基化的建立 (從頭開(kāi)始 DNA 甲基化),甲基化的維持和脫甲基。甲基化從寫入到擦除,牽涉著 DNMT 與 TET 兩大蛋白家族的“愛(ài)恨情仇”。
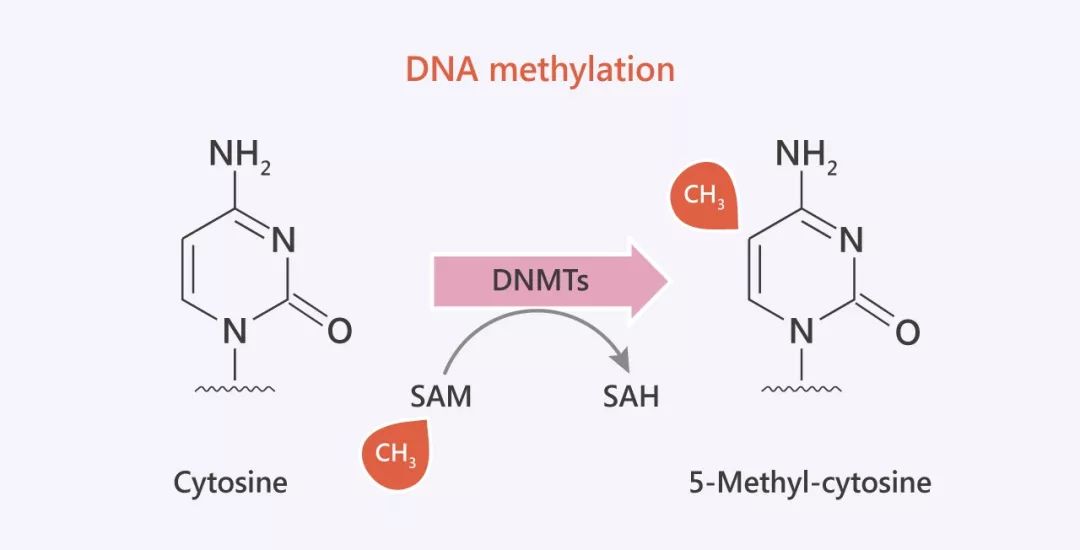
圖 2. 5mC 甲基化
DNMTs 家族蛋白又可細(xì)分為從頭甲基轉(zhuǎn)移酶 (DNMT3A, DNMT3B 和 DNMT3L) 和維持甲基轉(zhuǎn)移酶 (DNMT1) 兩大派別,在甲基化中發(fā)揮不同的作用。從頭甲基化:(De novo of DNA methylation )
DNA 甲基化通過(guò)稱為 DNMT3A 和 DNMT3B “從頭甲基化酶”在胚胎發(fā)育過(guò)程中建立。DNMT3A 和 DNMT3B 是對(duì)未甲基化的 DNA 具有活性的從頭甲基轉(zhuǎn)移酶,他們就像是甲基化歷程的“創(chuàng)業(yè)者”,不偏好半甲基化的 CpG 位點(diǎn),“從頭”甲基化先前未被甲基化的 DNA。
DNMT3 家族還存在一個(gè)成員:DNMT3L。與 DNMT3A/3B 相比,缺乏甲基基序和催化結(jié)構(gòu)域,但可以與 DNMT3A/3B 結(jié)合,刺激 DNMT3A/3B 的酶活性,在甲基化進(jìn)程中推波助瀾。
維持甲基化:(Maintenance of DNA methylation)
維持甲基化,就是雙鏈 DNA 的一條鏈已經(jīng)發(fā)生甲基化,已經(jīng)發(fā)生甲基化的鏈想將這種甲基化狀態(tài)“薪火相傳”(通過(guò)半保留復(fù)制)。為完成這一步驟,“維持甲基轉(zhuǎn)移酶” (DNMT1) 上線,DNMT1 更像是個(gè)獵頭,只認(rèn)識(shí)甲基化 CpG,沒(méi)有甲基化的 CpGs 不會(huì)成為維持甲基轉(zhuǎn)移酶 DMNT1 的底物。在這個(gè)進(jìn)程中,DNMT1 相互作用蛋白 UHRF1 (多結(jié)構(gòu)域蛋白 E3 泛素蛋白連接酶) 至關(guān)重要,因?yàn)?UHRF1 的 SET 和 RING 相關(guān) (SRA) 域?qū)Π爰谆?DNA 具有強(qiáng)烈的偏好,能幫助 DNMT1 在復(fù)制叉處靶向底物 CpG 二核苷酸。
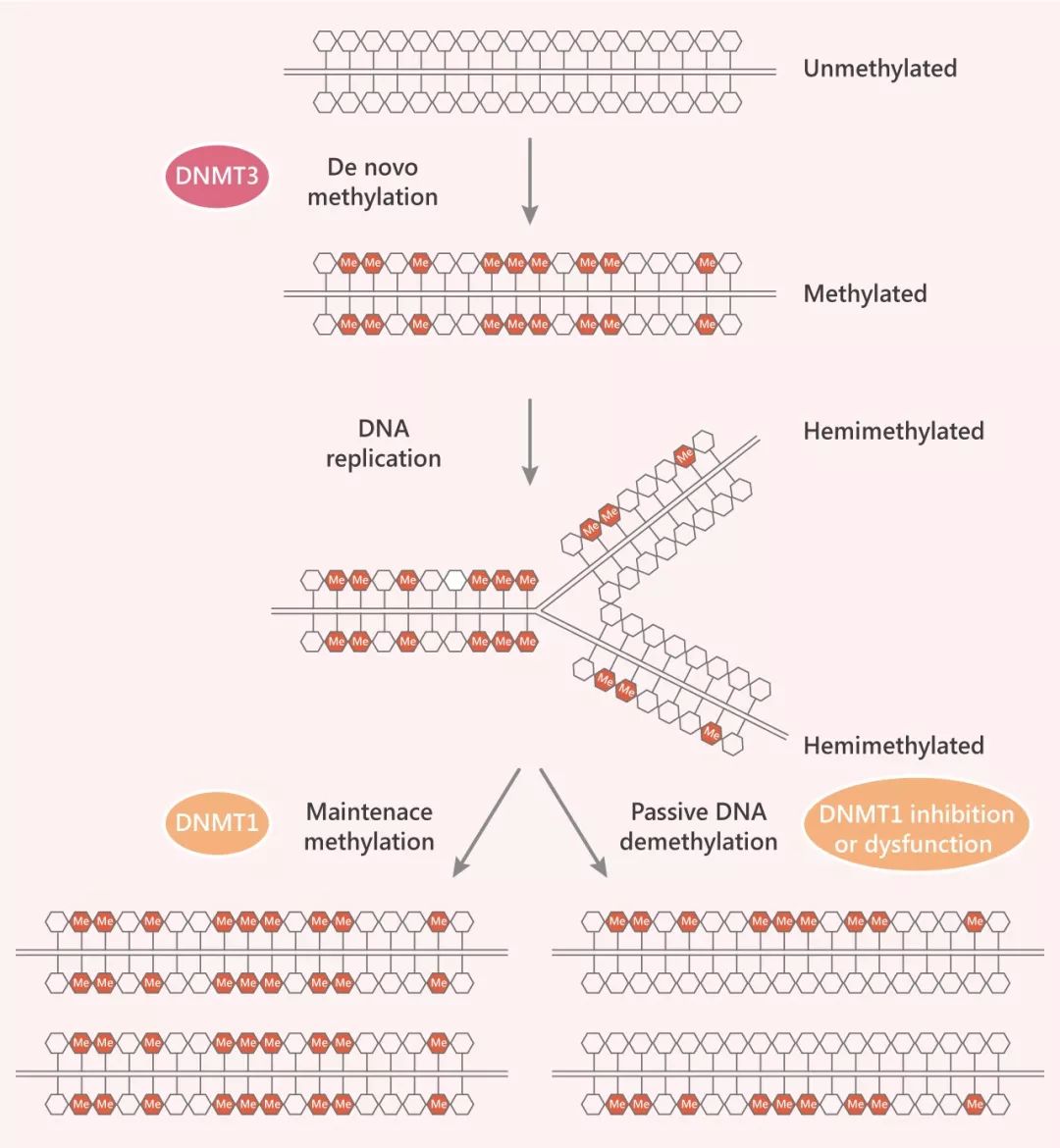
Tip 3:為確保親代模式在子代鏈上的復(fù)制,一種“維持甲基轉(zhuǎn)移酶” DNMT1,只會(huì)將甲基化 CpGs 堿基與甲基化的親代 CpG 配對(duì),非甲基化 CpGs 不會(huì)成為其底物。這種簡(jiǎn)單機(jī)制的結(jié)果是,DNA 甲基化模式會(huì)像 DNA 自身的堿基序列一樣被半復(fù)制。
除 DNA 甲基化——擦除蛋白 -Tet 啟動(dòng)的 DNA 脫甲基途徑
DNA 脫甲基涉及被動(dòng)和主動(dòng)脫甲基的兩個(gè)過(guò)程來(lái)實(shí)現(xiàn):5- 甲基胞嘧啶 (5mC) 作為 DNA 修飾的主要形式。結(jié)構(gòu)上,甲基通過(guò)牢靠的碳-碳雙鍵與胞嘧啶堿基的 5 位連接,為直接去除甲基加了一層保險(xiǎn)。機(jī)制上,前文提到的 DNMT3 以及官能伙伴 UHRF1 一起協(xié)助維持這種狀態(tài)。盡管如此,5mC 依然有自己的辦法逃脫層層枷鎖。
被動(dòng) DNA 脫甲基化:
功能性的 DNA 甲基化維持機(jī)制缺乏,使得 DNA 復(fù)制過(guò)程中 5mC 的逐步‘稀釋’ (passive solution),這個(gè)過(guò)程通常被稱為被動(dòng) DNA 脫甲基化 (參照 Fig 2)。該事件主要發(fā)生在分裂細(xì)胞中,DNMT1 的抑制或功能障礙會(huì)使新?lián)饺氲陌奏け3治醇谆瑵?rùn)物細(xì)無(wú)聲地降低了每個(gè)細(xì)胞分裂后的總體甲基化水平。主動(dòng) DNA 脫甲基化:
敲黑板嘍,該脫甲基化的過(guò)程,也稱為活性去甲基化。在這個(gè)過(guò)程中 TET 蛋白家族起到了舉足輕重的作用 (參照 Fig 3)。
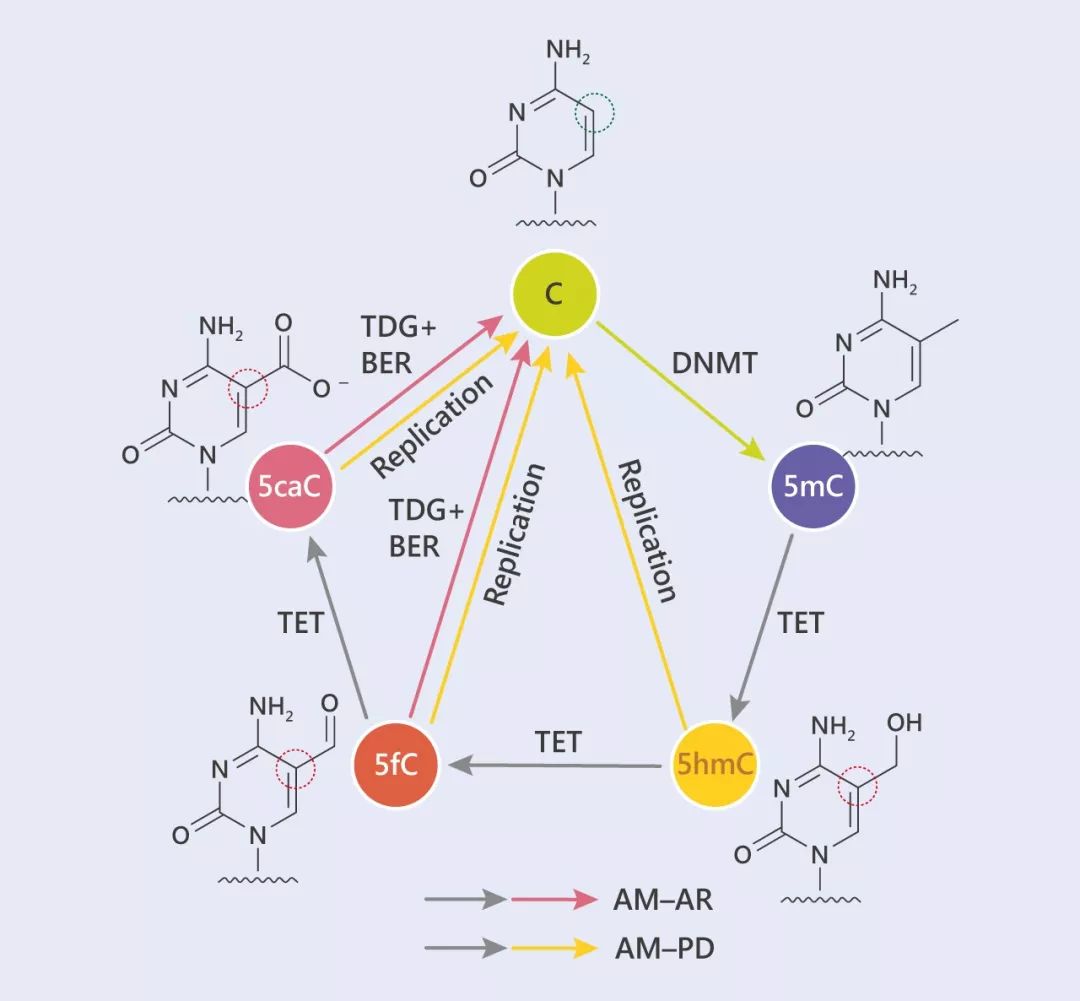 圖 4. Tet 啟動(dòng)的 DNA 脫甲基 (氧化位點(diǎn)已圈出)
圖 4. Tet 啟動(dòng)的 DNA 脫甲基 (氧化位點(diǎn)已圈出)在主動(dòng)脫甲基化的過(guò)程中,5mC 通過(guò) TET 蛋白,不斷被氧化,一步步去除“甲基化帽子”。
這個(gè)過(guò)程中。TET 蛋白像個(gè)探測(cè)器一樣,可以沿著 DNA “拉鏈”從一個(gè)點(diǎn)滑動(dòng)到另一個(gè) CpG 位點(diǎn),最神奇的是,TET 會(huì)優(yōu)先識(shí)別并且氧化同一 DNA 分子上的其他 CpG 位點(diǎn)。
TET 蛋白可以介導(dǎo) 5mC 迭代氧化為 5- 羥甲基胞嘧啶 (5hmC),5- 甲酰基胞嘧啶 (5fC) 和 5- 羧胞嘧啶 (5caC)。氧化的 5mC 衍生物不能用作 DNMT1 的底物,在復(fù)制的過(guò)程中被動(dòng)脫甲基作用而丟失,胸腺嘧啶DNA糖基化酶 (TDG) 介導(dǎo)的 5fC 和 5caC 切除的復(fù)制依賴稀釋,再加上堿基切除修復(fù) (BER),導(dǎo)致去甲基化,這個(gè)過(guò)程被定義為主動(dòng)修飾-主動(dòng)去除 (AM-AR)。此時(shí)的 5fC 和 5caC,就像是無(wú)業(yè)游民,TDG 能識(shí)別這種不正常的堿基并切除它們,隨后,AP 位點(diǎn)形成 (無(wú)嘌呤或者無(wú)嘧啶位點(diǎn)),開(kāi)始?jí)A基切除修復(fù)。另外,也可以通復(fù)制依賴 5hmC,5fC 或 5caC 的方式稀釋 DNA 甲基化,即主動(dòng)-被動(dòng)去除 (AM-PD)。
Tip 4: TDG :能夠識(shí)別不正常的堿基。如胞嘧啶 5mC 在脫氨基之后會(huì)直接形成尿嘧啶 5hmc,對(duì)于這些不正常的尿嘧啶,就可以被 TDG 切除掉,形成無(wú)嘌呤或者無(wú)嘧啶的位點(diǎn)。Base-excision repair:AP 位點(diǎn)形成,AP 核酸內(nèi)切酶在 AP 位點(diǎn)將 DNA 鏈切開(kāi),然后核酸外切酶 1 將包括 AP 位點(diǎn)在內(nèi)的 DNA 鏈切除。這種單個(gè)堿基的修復(fù)叫做堿基切除修復(fù)。
如果將甲基化的 DNA 比作是一匹千里馬的話,那么三個(gè)識(shí)別的蛋白家族 (MBD 蛋白質(zhì),UHRF 蛋白質(zhì)和鋅指蛋白) 算得上是識(shí)別它的伯樂(lè)了。
MBD 家族由 MeCP2 和 MBD1-6 組成,共享一個(gè)保守的 MBD 域,這也與甲基化 DNA 結(jié)合所必需的。MeCP2,MBD1 和 MBD2 還包含一個(gè) TRD 域,可幫助他們募集染色質(zhì)重塑核心加壓因子 (chromatin remodeling corepressors),從而引起轉(zhuǎn)錄沉默。如 MeCP2 的 TRD 結(jié)構(gòu)通過(guò)募集包含 Sin3A 共阻遏物和組蛋白脫乙酰基酶 (HDAC1 和 HDAC2) 的染色質(zhì)重塑復(fù)合體來(lái)參與介導(dǎo)基因沉默。
UHRF 蛋白 (如 UHRF1 和 UHRF2 ),前面在維持甲基化中已經(jīng)提過(guò)它的作用了。鋅指蛋白 (Kaiso,ZBTB4 和 ZBTB38) 則是通過(guò)鋅指結(jié)構(gòu)域與甲基化 DNA 結(jié)合。其中 Kaiso 被認(rèn)為是轉(zhuǎn)錄阻遏物,可以與活躍表達(dá)基因的未甲基化區(qū)域結(jié)合。DNA 甲基化與組蛋白修飾
DNA 甲基化和組蛋白修飾的關(guān)系千絲萬(wàn)縷。簡(jiǎn)而言之,DNA 甲基化有助于引導(dǎo)組蛋白修飾,組蛋白修飾又會(huì)指導(dǎo) DNA 甲基化。例如 DNMT3A 的 PWWP 域會(huì)與 H3K36me3 相互作用,增強(qiáng) DNMT3A 的活性。同樣,組蛋白修飾在 DNA 甲基化機(jī)制中也起著重要作用。拿 DNMT1 啟動(dòng)子過(guò)程中的甲基化來(lái)說(shuō),H3K4me3 可抑制DNMT3 酶。而在沒(méi)有 H3K4 甲基化的情況下,抑制會(huì)得以解除,從而使 DNA 甲基化。
H3K4me3 標(biāo)記了活躍的啟動(dòng)子,阻止了 DNMT3A 和 DNMT3B (以及DNMT3L) 的 ADD 域的染色質(zhì)結(jié)合,從而使 ADD 與甲基轉(zhuǎn)移酶 (MTase) 結(jié)構(gòu)域結(jié)合并自動(dòng)抑制 DNMT3 酶。在沒(méi)有 H3K4 甲基化的情況下,ADD 域與 H3K4 結(jié)合,并且自動(dòng)抑制得以解除,從而 MTase 域可以使 DNA 甲基化。
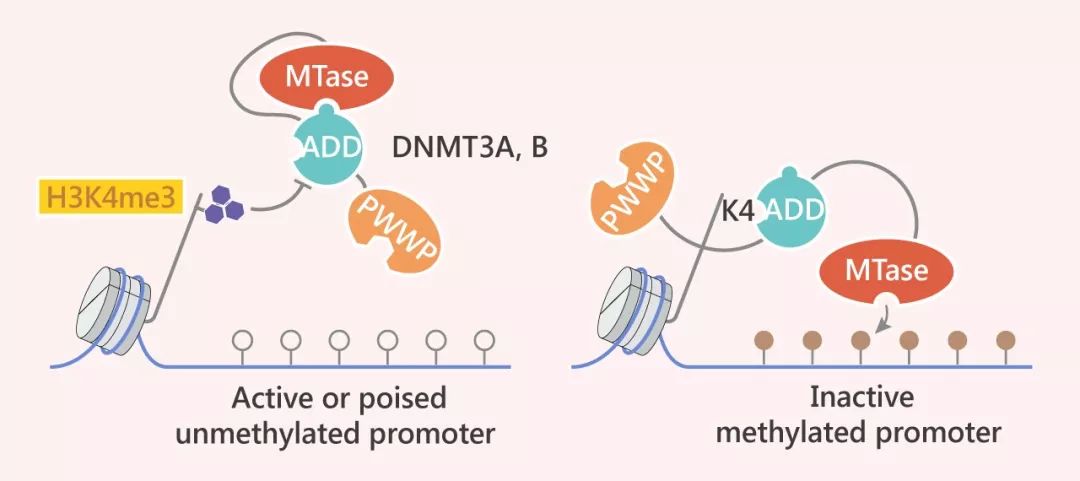
Tip 5:H3K4me3 標(biāo)記了活躍的啟動(dòng)子,阻止了 DNMT3A 和 DNMT3B (以及DNMT3L) 的ADD域的染色質(zhì)結(jié)合,從而使ADD 與甲基轉(zhuǎn)移酶 (MTase)結(jié)構(gòu)域結(jié)合并自動(dòng)抑制 DNMT3 酶。在沒(méi)有H3K4甲基化的情況下,ADD 域與 H3K4 結(jié)合,并且自動(dòng)抑制得以解除,從而 MTase 域可以使 DNA 甲基化。
在高等動(dòng)物中,癌細(xì)胞中的 DNA 甲基化模式明顯“扭曲”。大約 70% 的基因擁有與啟動(dòng)子相關(guān)的 CpG 島 (CG 堿基頻率高的區(qū)域),而維持未甲基化的啟動(dòng)子 CpG 島對(duì)提高轉(zhuǎn)錄潛能有積極作用,并與活性的組蛋白修飾如組蛋白 H3 和 H4 的乙酰化和 H3K4 的甲基化有關(guān)。
相反,癌細(xì)胞明顯表現(xiàn)出 CpG 島的密集高甲基化,包括基因間區(qū)和重復(fù)元件在內(nèi)的大量染色質(zhì)的低甲基化。密集甲基化的 CpG 島能夠讓原本松散的染色質(zhì)緊縮在一起,抑制與“抑制性修飾” (H3K9me2/3,H3K27me3 和/或 H4K20me3) 相關(guān)的基因表達(dá),可見(jiàn) DNA 甲基化顯著著影響癌細(xì)胞的基因組格局。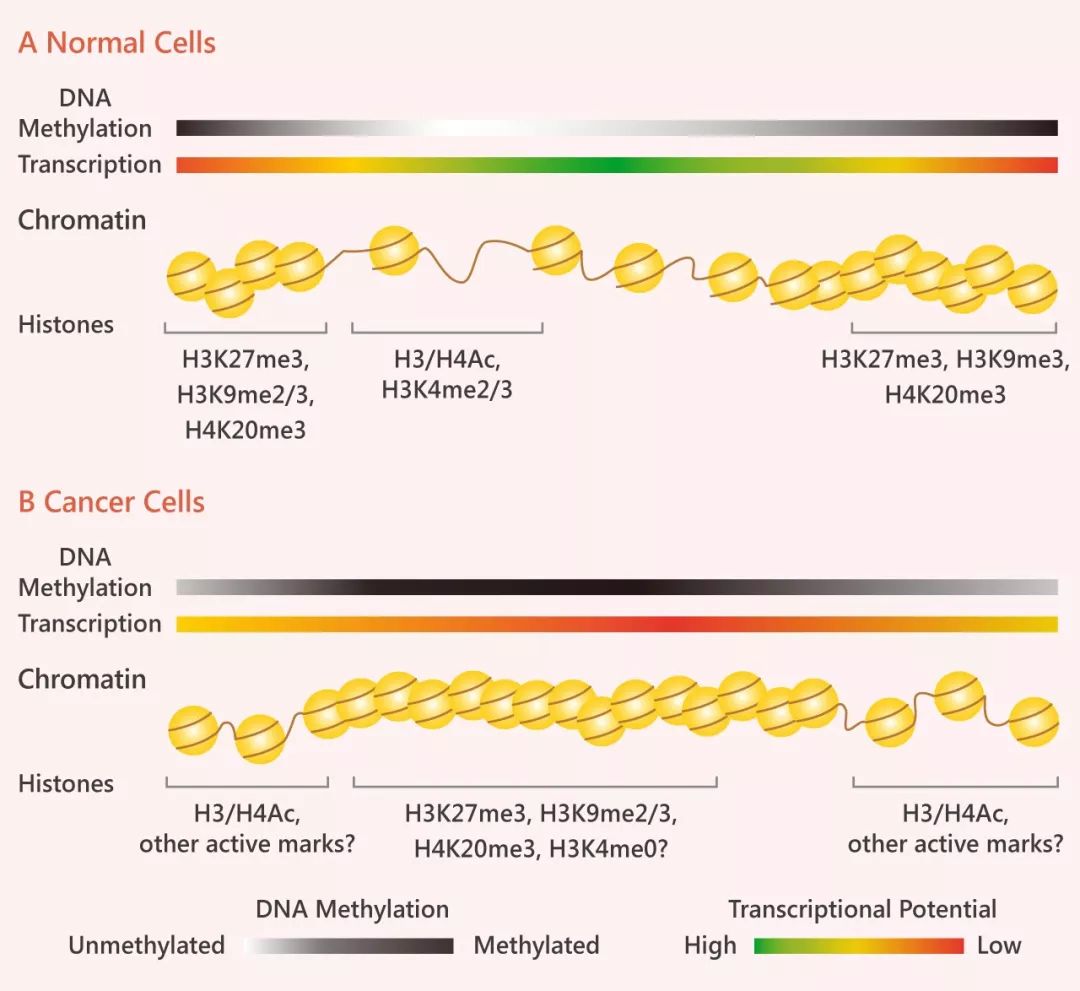
Tip 6: DNA 被包裹在組蛋白的八聚體中,形成核小體,即染色質(zhì)的最小單位。
正常情況下轉(zhuǎn)錄時(shí)是開(kāi)放松散的染色質(zhì)結(jié)構(gòu):組蛋白 H3 和 H4 的高度乙酰化,組氨酸 H3 在賴氨酸 4 (H3K4me2/2/3) 處的二甲基和三甲基化 (Fig 5A)。然而,當(dāng)轉(zhuǎn)錄由于被甲基化受阻時(shí),受阻區(qū)域染色質(zhì)結(jié)構(gòu)就會(huì)變得緊縮在一起,H3/H4 乙酰化和 H3K4 甲基化不顯示,取而代之的是富含 H3K9 (H3K9me2/3),H3K27的三甲基化 (H3K27me3) 和 H4K20 的三甲基化 (H4K20me3)(Fig 5)。
DNA 甲基化抑制劑
DNA 甲基化抑制劑一般分為核苷類似物和非核苷類似物化合物。核苷類似物如 Decitabine,5-Azacytidine,Zebularine,它們可摻入 DNA 中,摻入后,它們充當(dāng) DNMT 酶的自殺底物。非核苷化合物可以抑制 DNA 甲基轉(zhuǎn)移酶活性,如 RG108,DC-05,它們直接阻斷 DNA 甲基轉(zhuǎn)移酶的活性。Decitabine (NSC 127716)
Decitabine,地西他濱,F(xiàn)DA 批準(zhǔn)的 DNA 甲基轉(zhuǎn)移酶 (DNMTs) 抑制劑。通常用于治療骨髓增生異常綜合征 (MDS) 和急性髓性白血病 (AML)。Decitabine 具有雙重的劑量依賴性作用機(jī)制。高劑量時(shí),Decitabine 通過(guò)摻入 DNA 代替胞嘧啶后,可將 DNA 甲基轉(zhuǎn)移酶共價(jià)捕獲到 DNA 中,從而誘導(dǎo)細(xì)胞毒性作用。在較低劑量下其抗腫瘤效果可能是由于它能夠抑制 DNA 甲基化和重新激活腫瘤抑制基因。
5-Azacytidine (Azacitidine)
5-Azacytidine,阿扎胞苷,DNA 和 RNA 核苷胞苷類似物,也是FDA 批準(zhǔn)的 DNA 甲基轉(zhuǎn)移酶 (DNMTs) 抑制劑,5-Azacytidine 必須與 DNA 結(jié)合以共價(jià)捕獲 DNA 甲基轉(zhuǎn)移酶。5-Azacytidine 也可以摻入到 RNA 中。
RG108 (N-Phthalyl-L-tryptophan)
RG108,非核苷類似物,是一種靶向人 DNA 甲基轉(zhuǎn)移酶 (DNMTs) 的抑制劑,IC50 值為 115 nM。RG108 結(jié)合并阻斷 DNMTs 的活性位點(diǎn)。RG108 缺少與 Decitabine 相關(guān)的高水平的細(xì)胞毒性。RG108 可在不影響著絲粒衛(wèi)星序列甲基化狀態(tài)的情況下,引起腫瘤抑制基因的去甲基化和再激活。
參考文獻(xiàn):
[1] Zhang W, et al. DNA methyltransferases and their roles in tumorigenesis. Biomark Res. 2017Jan 20;5:1.[2] McCabe MT, et al. Cancer DNA methylation: molecular mechanisms and clinicalimplications. Clin Cancer Res. 2009 Jun 15;15(12):3927-37.
[3] Yu J,et al. DNA methyltransferase expression in triple-negative breast cancerpredicts sensitivity to decitabine.J Clin Invest. 2018 Jun 1;128(6):2376-2388.
[4] Issa JP,et al. Phase 1 study of low-dose prolonged exposure schedules ofthe hypomethylating agent 5-aza-2'-deoxycytidine (decitabine)in hematopoietic malignancies. Blood. 2004Mar 1;103(5):1635-40. Epub 2003 Nov 6.
[5] Christman JK. 5-Azacytidine and 5-aza-2'-deoxycytidine as inhibitors of DNAmethylation: mechanistic studies and their implications for cancer therapy. Oncogene. 2002Aug 12;21(35):5483-95.
[6] Qiu X, et al. Equitoxic doses of 5-azacytidine and 5-aza-2'deoxycytidine induce diverse immediate and overlapping heritable changes inthe transcriptome. PLoS One. 2010 Sep 29;5(9).
[7] Brueckner B,et al. Epigenetic reactivation of tumor suppressor genes by a novelsmall-molecule inhibitor of human DNA methyltransferases. Cancer Res. 2005 Jul 15;65(14):6305-11

- Amyloid β-Peptide的結(jié)構(gòu)、作用機(jī)制及在科研中的應(yīng)用
- PARP抑制劑Olaparib的作用機(jī)理及在科研中的應(yīng)用
- 磷脂聚乙二醇甘露糖(DSPE-PEG2000-Mannose)的結(jié)構(gòu)、特性與應(yīng)用
- 疫苗佐劑QS-21的作用機(jī)理及在疫苗研究中的應(yīng)用
- 生物指示劑最短培養(yǎng)時(shí)間(MIT)測(cè)定全新方案介紹
- ReadiLin抗體標(biāo)記試劑盒的優(yōu)勢(shì)及應(yīng)用場(chǎng)景
- MST 技術(shù)助力難純化蛋白的親和力分析
- 抑制劑BIIB021在細(xì)胞實(shí)驗(yàn)和動(dòng)物實(shí)驗(yàn)的溶解方案
- 西美杰成功參加第二十四屆中國(guó)生物制品大會(huì)
- 2025AACR年會(huì)圓滿落幕,MCE攜試劑產(chǎn)品亮相芝加哥
- 西美杰成功參加第十屆生物制藥穩(wěn)定性論壇
- 投稿有禮-2025 MCE中國(guó)生命科學(xué)研究促進(jìn)獎(jiǎng)?wù)絾?dòng)
- 艾偉拓&安拓思會(huì)議邀請(qǐng):集采時(shí)代的復(fù)雜制劑新方向
- 開(kāi)學(xué)季,科研福利到!本生生物提取試劑盒限時(shí)特惠
- 西安百螢生物酪胺染料限時(shí)促銷,購(gòu)買即免費(fèi)送電影票
- 美德聲推出通過(guò)LC-MS/MS擴(kuò)大新生兒篩查的檢測(cè)試劑