轉錄因子Sp9參與神經紋狀體蒼白球發育過程
轉錄因子Sp9參與神經紋狀體蒼白球發育過程
研究人員發現鋅指轉錄因子——Sp9,在LGE祖細胞中廣泛表達,對維持有絲分裂期后的紋狀體蒼白球MSNs至關重要,為我們理解神經元發育過程提供了新的證據。研究背景
紋狀體是基地神經節的重要組成部分,是一類中型多棘神經元(MSNs)。MSNs的兩個重要的基地神經節亞型分別是紋狀體黑質(直接通路)和紋狀體蒼白球(間接通路)。紋狀體中5%-10%的神經細胞為多棘內在神經元。已有研究發現一些轉錄因子(TFs)參與紋狀體黑質MSNs的形成,但是TFs在紋狀體蒼白球MSNs形成過程中的作用還不清晰。本研究通過對一個鋅指蛋白轉錄因子Sp9的深入研究,闡述了該轉錄因子在紋狀體蒼白球發育過程中的重要作用研究思路
研究結果
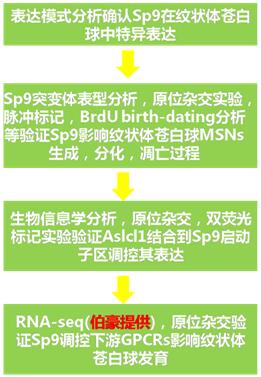
1. Sp9表達模式分析表明其在神經紋狀體蒼白球中特異表達
為了系統性了解的轉錄因子Sp9在大腦中表達和功能,研究人員構建了Sp9多克隆抗體和突變體alleles,Sp9-LacZ null allele(Sp9LacZ/+)和Sp9 floxed allele(Sp9Flox/+)。檢測顯示,Sp9-LacZ在E10.5期的神經節(GEs)中廣泛表達。免疫細胞化學實驗和RNA雜交實驗表明Sp9 RNA和蛋白在E13.5期的室下區域(SVE)和地幔區域的LGE,MGE和CGE區域廣泛表達。同時,在SVE的Sp9+細胞中,神經前體蛋白Ascl1和細胞增殖marker Ki67也有明顯表達。在胚胎發育階段,Sp9在大部分的遷移皮質神經節中表達。
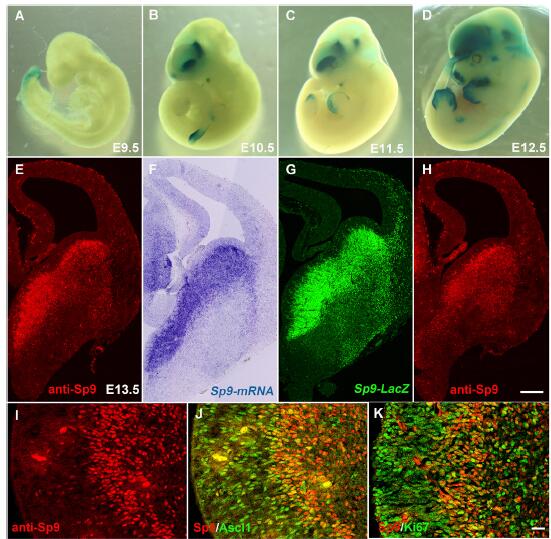
通過構建Sp9-Cre knockin小鼠,研究人員發現Sp9+祖細胞產生了96%以上的皮層中間神經元和VIP(腸血管活性肽)+亞型和幾乎所有的紋狀體中間神經元。Foxp1是特異性的在有時分裂后期的MSNs細胞中表達,結果顯示所有Foxp1+細胞中都有Sp9-Cre表達,表明Sp9-Cre在紋狀體黑質和紋狀體蒼白球MSNs中被激活。此外,研究人員也發現Sp9-Cre在嗅球細胞(OB)中間神經元中表達。
通過Drd2-EGFP轉基因小鼠,研究人員構建了Sp9紋狀體表達模型小鼠。結果顯示,Sp9在胎兒紋狀體中強烈表達,一直持續到嬰兒時期。在E16.5,P0, P5,P17和P35期的紋狀體蒼白球中都表達Sp9。該結果表明,在MSNs有絲分裂期后,Sp9特異性地在紋狀體蒼白球MSNs中特異表達。
2. Sp9突變小鼠中紋狀體蒼白球MSNs的生成和凋亡受影響
為探究Sp9的生物學功能,研究人員對Sp9 Laz/Laz突變小鼠進行了分析。結果顯示,突變體小鼠發育較弱,持續到P7還是發育無力,在P14開始死亡,在P22期已沒有存活的小鼠。同時,Sp9突變體小鼠的大腦大小和重量也都比正常的小。
表型分析顯示,Sp9突變體小鼠大腦紋狀體嚴重萎縮,在P9期體積是對照的54%。同時,相對于Sp9LacZ/+小鼠,Drd2-EGFP 小鼠中Foxp1+細胞減少57%,Sp9LacZ/LacZ小鼠中,Foxp1+/Drd2-GFP+細胞減少了97%,且在紋狀體中非均勻分布。在背內側紋狀體中,Foxp1+/Drd2-GFP+細胞也很難檢測到,但出現在了側紋狀體中。對Drd2的原位雜交實驗也確認了該類細胞的缺失。此外,突變體小鼠的Enk+細胞嚴重減少,進一步表明紋狀體蒼白球MSNs的缺失。只有很少的Drd2-GFP+細胞出現在背側紋狀體中,屬于中間神經元亞型。另外,Drd1-GFP在紋狀體特異性的在黑質細胞中表達,Sp9LacZ/LacZ突變體中紋狀體黑質MSNs的GFP表達并沒有明顯變化。該結果表明,Sp9突變只影響了紋狀體蒼白球的發育。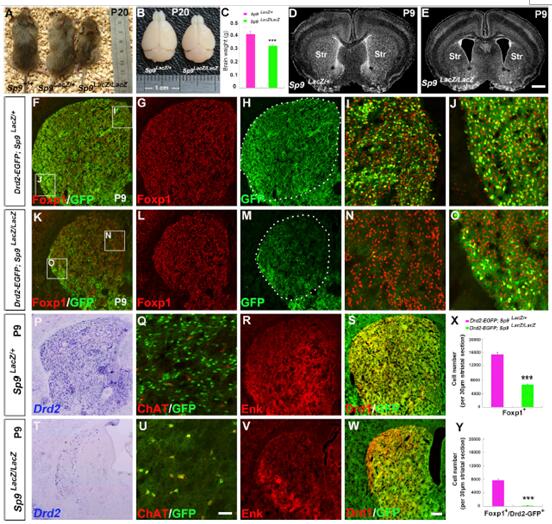
研究人員對E13.5和E15.5小鼠LGE進行30-min BrdU 脈沖標記。結果顯示,E13.5期突變體BrdU+細胞在SVZ中減少。E15.5期突變體中BrdU+細胞在SVZ祖細胞和SVZ Foxp1+有絲分裂期后MSNs中都減少。
研究人員接下來進行了BrdU birth-dating分析。在E12.5期注射BrdU,在突變體和對照P0期統計BrdU+和BrdU+/Foxp1+細胞,發現兩者都明顯減少。因此,缺失了Sp9影響了LGE SVZ的循環祖細胞的缺失。
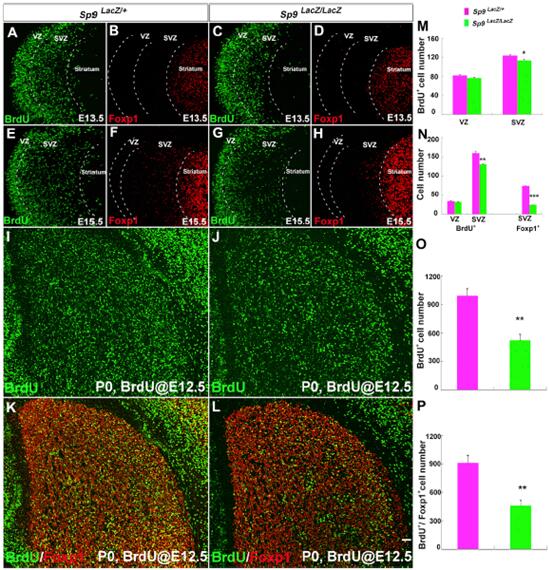
對Sp9LacZ突變體中β-gal+和β-gal+/Foxp1+細胞數目統計,發現數量顯著減少。由于Sp9主要在紋狀體蒼白球細胞中表達。所以,Sp9的缺失影響了紋狀體蒼白球MSNs的產生。
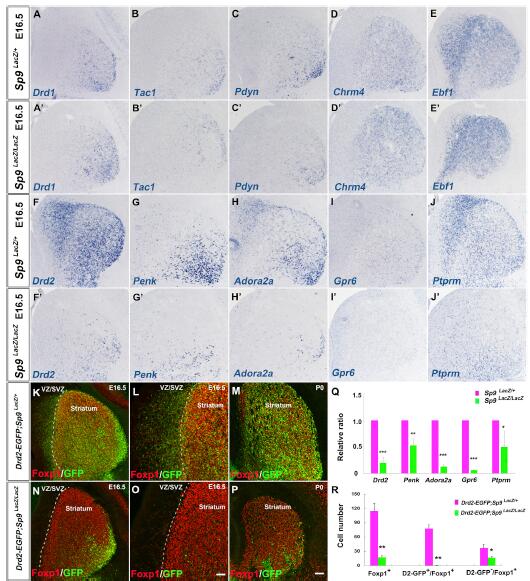
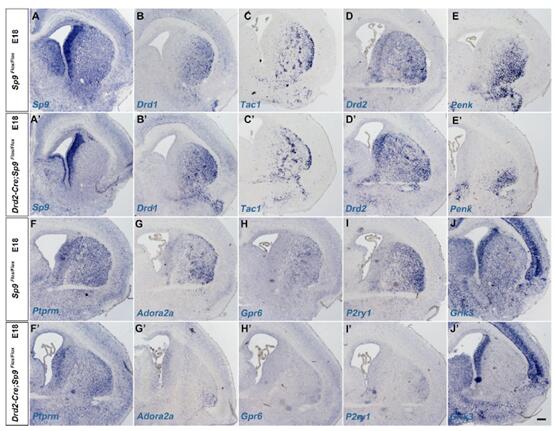
對E16.5期的紋狀體MSNs中10個紋狀體MSN markers進行原位RNA雜交檢測,發現5個紋狀體黑質marker(Drd1,Tac1,Ebf1,Pdyn,Chrm4)和5個紋狀體蒼白球marker(Drd2, Penk, Gpr6, Adora2a, Ptprm)在Sp9LacZ/+controls中都表達。在Sp9 LacZ/LacZ突變體SVZ和紋狀體中,紋狀體蒼白球MSN markers顯著下調,qRT-PCR結果也驗證了該結果。但是,紋狀體黑質markers表達沒有顯著影響。
對E16.5和P0期紋狀體Drd2-EGFP檢測,發現在突變體的SVE和紋狀體中,Foxp1+/Drd2-GFP+ MSD細胞更少。因此,這些結果都顯示,Sp9特異性地促進紋狀體蒼白球MSNs的生成。
研究人員接下來分析了Caspase-3表達,以檢測出生后Sp9LacZ/LacZ突變鼠紋狀體的細胞死亡。結果顯示,突變體在出生后不同時間Caspase-3+細胞增加顯著大于對照。由于在Caspase-3+細胞中并沒有檢測到Drd2-GFP的表達和紋狀體黑質MSNs的減少,所有在Sp9突變體紋狀體中,死亡的細胞為未成熟的紋狀體蒼白球MSNs。
為探究突變體紋狀體蒼白球 MSNs凋亡是否為Bcl-2-associated X 蛋白(Bax)-依賴,研究人員構建了Sp9LacZ/LacZ; Bax-/-雙突變體。結果顯示,雙突變體在P3期紋狀體細胞死亡幾乎消失,Foxp1+細胞數量明顯高于單突。值得注意的是,這種增加的Foxp1+細胞并不能轉化成成熟的紋狀體蒼白球MSNs。因此,除了促進LGE增殖和生成,Sp9還影響了紋狀體蒼白球神經元的分化和成熟過程。
為進一步分析Sp9對紋狀體蒼白球MSNs有絲分裂期后的作用,研究人員構建了條件性敲除突變系Drd2-Cre;Sp9Flox/Flox;Rosa-YFP+小鼠。突變體中的Foxp1+和Foxp1+/GFP+細胞明顯減少。在P0,P3和P5期條件性突變體Caspase-3+細胞數量也明顯上升。因此,Sp9條件性缺失突變體的Drd2+紋狀體蒼白球MSNs的缺失與細胞程序性死亡相關,說明Sp9對于紋狀體蒼白球MSNs有絲分裂期后的生存相關。
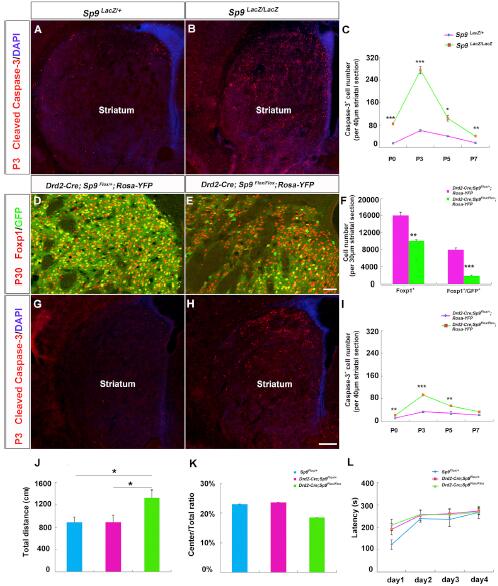
那么,這種紋狀體蒼白球MSNs的缺失會造成什么樣的生理問題呢?結果顯示,條件性突變小鼠的移動能力受到了影響,表現出更強的移動能力。但是在旋轉測試能力中并沒有顯著差異。因此,Drd2-Cre;Sp9Flox/Flox小鼠的快速移動能力增強,但不影響焦慮和運動協調能力。
3. Sp9作用分子機制探究
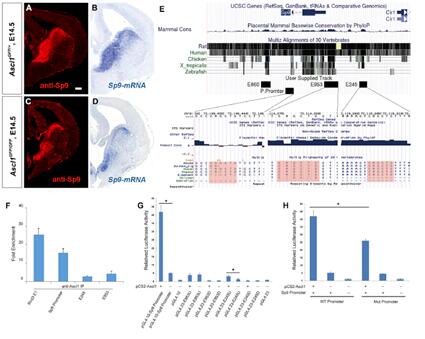
已有報道顯示Ascl1與小鼠Dre2+MSNs的生成有關。研究人員用原位RNA雜交進行了確認。與對照相比,Ascl1GFP/GFP突變小鼠中在P0期Drd2表達顯著下降。在E14.5和E16.5期,Ascl1GFP/GFP突變小鼠背側LGE SVE中的SP9蛋白與RNA表達都減少,暗示著Ascl1可以正向調控了Sp9的表達。
為進一步驗證其調控關系,研究人員分析了Sp9的啟動子區域,發現有Ascl1的保守結合位點(E-box sites, CAGCTG or CACCTG)。ChIP qPCR實驗表明兩者確實能夠結合。
對P19期胎瘤細胞進行雙熒光轉錄活性實驗也驗證了Ascl1可以通過(CAGCTG)結合Sp9 E245激活Sp9的轉錄。因此,這些結果表明Ascl1通過直接結合到Sp9的啟動子區促進LGE SVE中Sp9的表達。
研究結論
該研究通過表達模式分析發現了轉錄因子Sp9在LEG祖細胞中的廣泛表達。運用Sp9-null突變小鼠,發現Sp9對于紋狀體蒼白球MSNs的增殖,凋亡密切相關。進一步的ChIP qPCR實驗驗證了Sp9受到上游Ascl1的調控。RNA-seq實驗表明Sp9促進了下游GPCRsa家族蛋白的表達實現生物學功能。該研究深入理解神經節發育過程中紋狀體蒼白球MSNs的產生,分化和生存過程提供了新的證據。
原文出處: Zhang Q, Zhang Y, Wang C, et al. The Zinc Finger Transcription Factor Sp9 Is Required for the Development of Striatopallidal Projection Neurons. Cell Rep. 2016 ,16(5):1431-44.